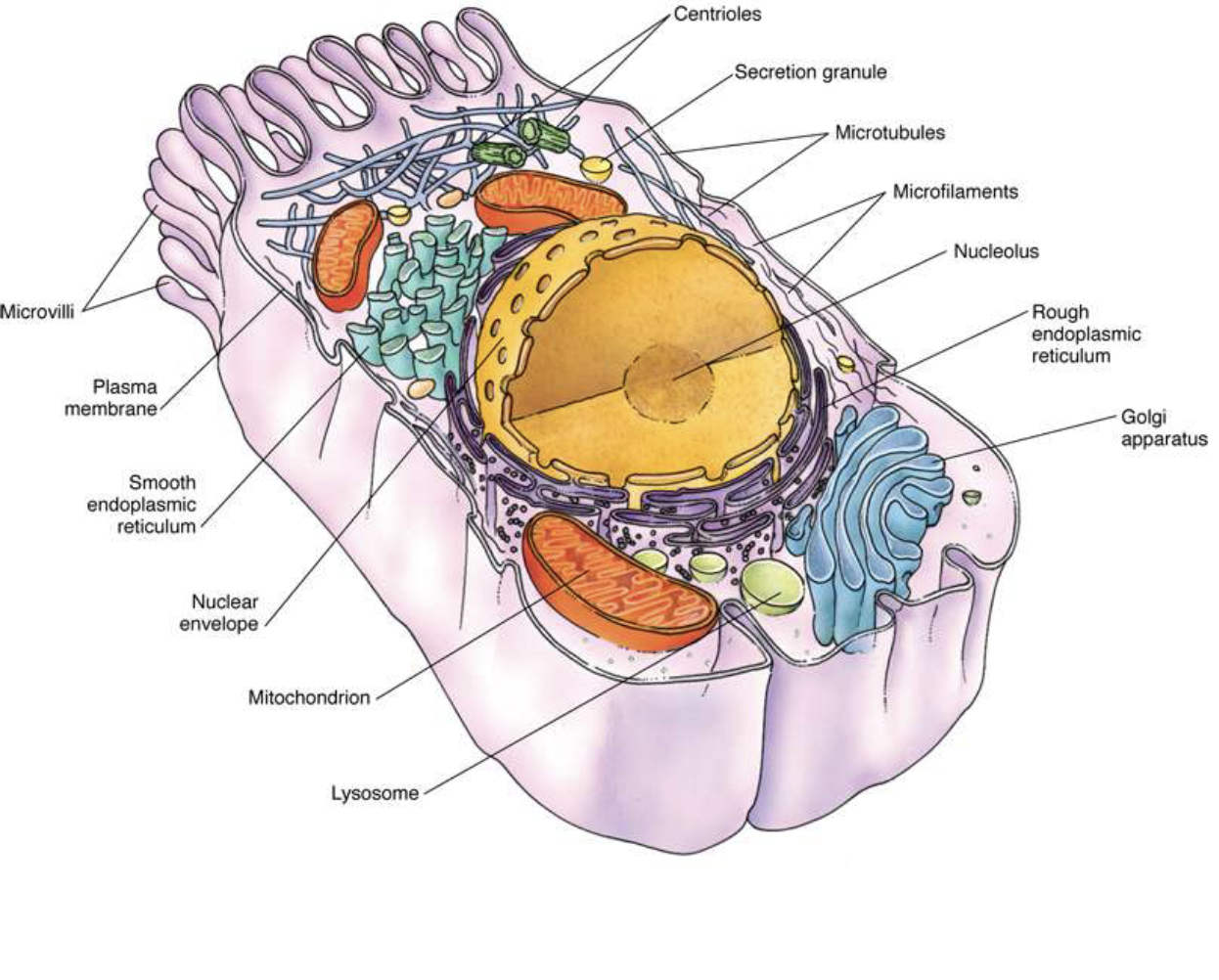
A cell is subdivided into two compartments:
- cytoplasm, extending from the plasma membrane to the nuclear envelope
- and karyoplasm (nucleoplasm), the substance forming the contents of the nucleus.
The bulk of the cytoplasm is water, in which various inorganic and organic chemicals are dissolved and/or suspended.
This fluid suspension, the cytosol, contains organelles, metabolically active structures that perform distinctive functions.
Cell Membrane
A fluid mosaic model of the cell membrane.
Each cell is bounded by a cell membrane (also known as the plasma membrane) that functions in:
- Maintaining the structural integrity of the cell
- Controlling movements of substances in and out of the cell (selective permeability)
- Regulating cell–cell interactions
- Recognition, via receptors, antigens, and foreign cells as well as altered cells
- Acting as an interface between the cytoplasm and the external milieu
- Establishing transport systems for specific molecules
- Transducing extracellular physical or chemical signals into intracellular events.
Endocytosis and the Endosomal System of Vesicles
The process whereby a cell ingests macromolecules and other substances from the extracellular space is referred to as endocytosis.
The endocytosed material is engulfed in a vesicle appropriate for its volume.
If the vesicle is large (>250 nm in diameter), the method is called phagocytosis (cell eating) and the vesicle is a phagosome.
The endosomal pathways. CURL, compartment for uncoupling of receptor and ligand.
If the vesicle is small (<150 nm in diameter), the type of endocytosis iscalled pinocytosis (cell drinking) and the vesicle is a pinocytotic vesicle.
Shortly after their formation, pinocytotic vesicles lose their clathrin coats (which return to the pool of clathrin in the cytosol) and fuse with early endosomes, a system of vesicles and tubules located near the plasma membrane.
The endosomal pathways. CURL, compartment for uncoupling of receptor and ligand.
If the entire contents of the pinocytotic vesicle requires degradation, the material from the early endosome is transferred to a late endosome.
This similar set of tubules and vesicles, located deeper in the cytoplasm near the Golgi apparatus, helps to prepare its contents for eventual destruction by lysosomes.
The endosomal pathways. CURL, compartment for uncoupling of receptor and ligand.
Cytoskeleton
REMEMBER THE SIZE OF THE FILAMENTS.
The shapes of cells, their ability to move, and the intracellular pathways within cells are maintained by a system of tubules and filaments known as the cytoskeleton.
Three dimensional network of protein distributed throughout the cytoplasm with roles:
- Cell movement (blood cells along the blood vessels walls, migration of fibroblasts during wound healing and in the embryonic development)
- Support and strength for the cell
- Phagocytosis
- Cytokinesis
- Cell-cell and cell-extracellular matrix adherence
- Change in cell shape
The component of the cytoskeleton were originally identified by electron microscopy as cytoplasmic “cables” that fell into 3 size groups.
The cytoskeleton has three components:
- Microtubules
- Thin filaments or Microfilaments
- Intermediate filaments
The only organelle that is composed of cytoskeletal elements is the centriole, a structure assembled from microtubules.
Microfilaments or Thin Filaments – (thick)
Thin Filaments are composed of actin monomers: two chains of its glomerular form G-actin, coiled around each other to form a filamentous protein: F-actin.
F-actin is a versatile and abundant cytoskeletal component forming static and contractile bundles. It anchors to the plasma membrane, thus it contributes to the shape of the cells.
Actin filaments are polar.
Thin filaments are 6-7 nm thick and possess a faster growing plus end and a slower-growing minus end.
Capping proteins attach to the plus end, terminating the lengthening of the filaments.
In cell migration, actin filaments undergo treadmilling to extend the leading edge, while interacting with non-muscle myosin to provide the contractile force necessary for movement.
Treadmilling
Treadmilling refers to the dynamic process by which actin monomers are added to one end of a filament (the “plus end”) while simultaneously being removed from the other end (the “minus end”).
This results in the overall movement of the filament towards the plus end while maintaining a constant length.
The minus ends (pointed end) grow less rapidly than the plus ends (barbed end) of actin filaments.
gActin bound to ATP associates with the rapidly growing plus ends, and the ATP bound to actin is then hydrolyzed to ADP.
During treadmilling, the addition and removal of actin monomers are regulated by various proteins that bind to the filament ends.
- The protein profilin, binds G-actin and helps to add actin monomers to the plus end,
- While the protein cofilin, an actin depolymerazing factor helps to remove actin monomers from the minus end, this replenishes the pool of G-actin for subsequent network assembly.
- When the actin filament reaches the desired length, member of a family of capping proteins (such as gelsolin), attach to the plus-end, terminating the lengthening of the filament.
- The cell membrane poly-phospho-inositide removes the actin cap gelsolin, permitting elongation of the actin filament.
There are 3 classes of actin:
-
-actin → muscle
-
-actin and -actin → no muscle cells
-
Responsible for motility via association with different actin binding protein. The most commonly known is myosin.
-
Gelsolin is a cap protein, stabilizing the end of the filaments
-
Cofillin: induces depolymerization by the removal G-actin at the minus end.
Mobility: Filopodium and Lamellipodium
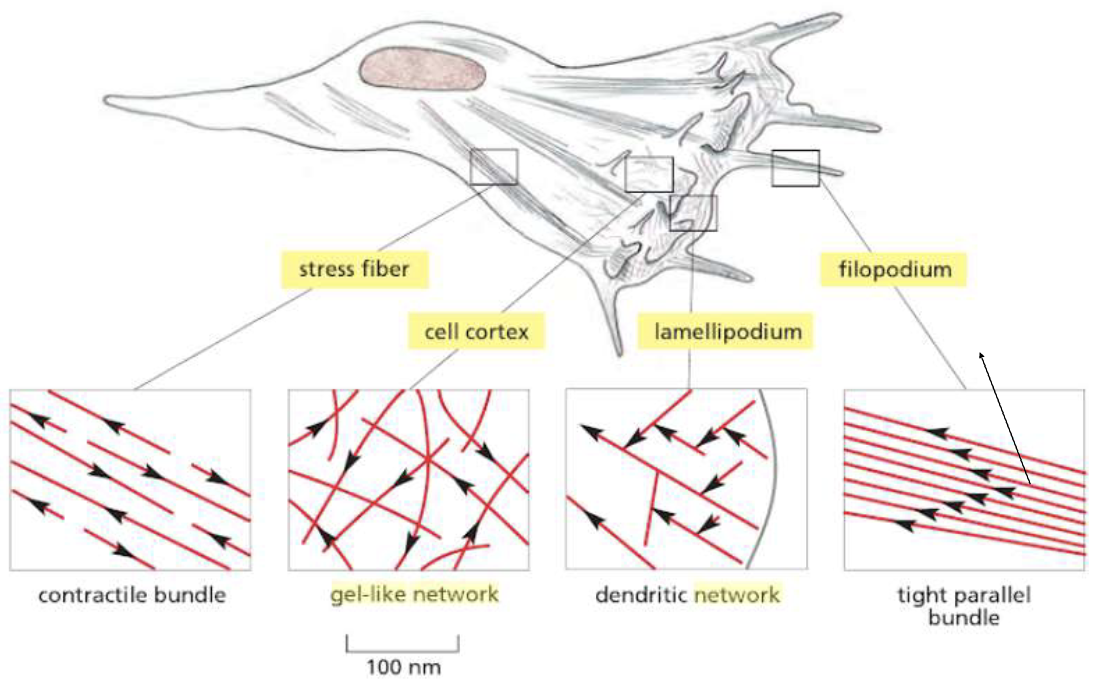
Thin filmaments can be found in specific projections in the front of a cell (when it wants to migrate) called lamellopodium and filopodium. (Branching F-Actin in the Leading Edge)
They help the cells move and gain nutrients by increasing the cell surface.
The cell cortex is made by filaments of actin arranged as a gel-like network.
The contractile bundles:
- Are associated with myosin
- The actin filaments are arranged loosely, parallel to each other, with plus and minus end alternating in direction
- Roles:
- Organelles and vesicles movements within the cells
- Endo and Exocytosis
- Contractile process and muscle contraction
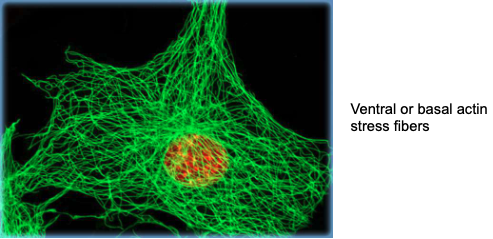
Spatial organization of actin stress fibers in adherent cells.
Super resolution images of normal cells stained with phalloidin highlights distinct stress fiber subtypes (color-coded):
- dorsal stress fibers,
- transverse arcs, (all together they help the cell to maintain the 3D structure)
- ventral stress fibers (are not connected to the periphery)
- and the perinuclear actin cap (top left).
→ they are holding and squeezing the nucleus in place.
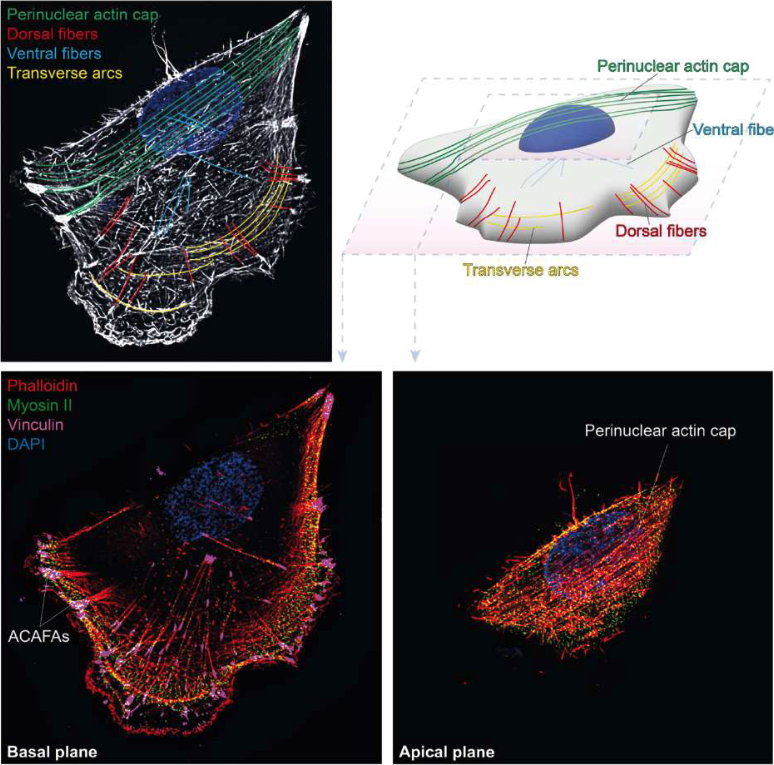
A schematic 3D representation of the same cell illustrates their spatial arrangement within the cell body (top right).
Basal (bottom left) and apical (bottom right) planes show actin together with myosin II and vinculin, highlighting actomyosin contractility and adhesion-associated structures, including actin cap–associated focal adhesions (ACAFAs).
Nuclei are counterstained with DAPI.
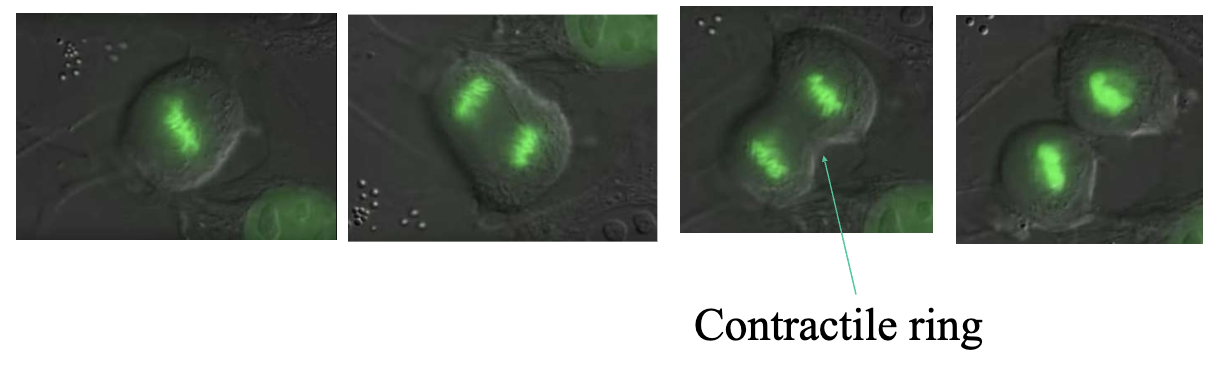
Contractile Ring
Microfilaments are also responsible for the formation of the contractile ring during mitotic division, made of F-Actin and myosin and it’s enough to help the cells divide.
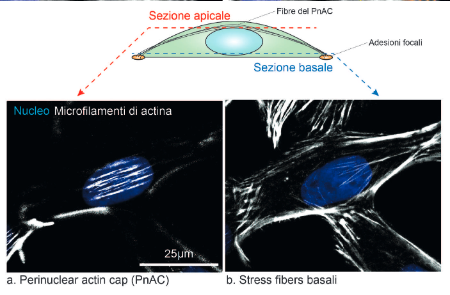
Spatial organization of actin stress fibers in adherent cells
The assembly and function of perinuclear actin cap in migrating cells
When the cell decides to move there’s the formation of the MIGRATION FRONT: there’s the formation of the lamellipodia and filopodia. They are moving in the direction of formation of the lamellipodia.
Polarized cells display a conical shape with predominantly convex leading edge and concave cell sides. The nucleus (blue) is located at the cell rear with longer nuclear axis aligned with the direction of cell migration (black arrow).
Actin forms different stress fibers that vary in their location and in the attachment to focal adhesions.
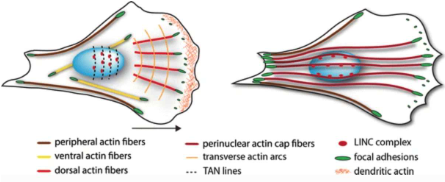
Actin cap known partners
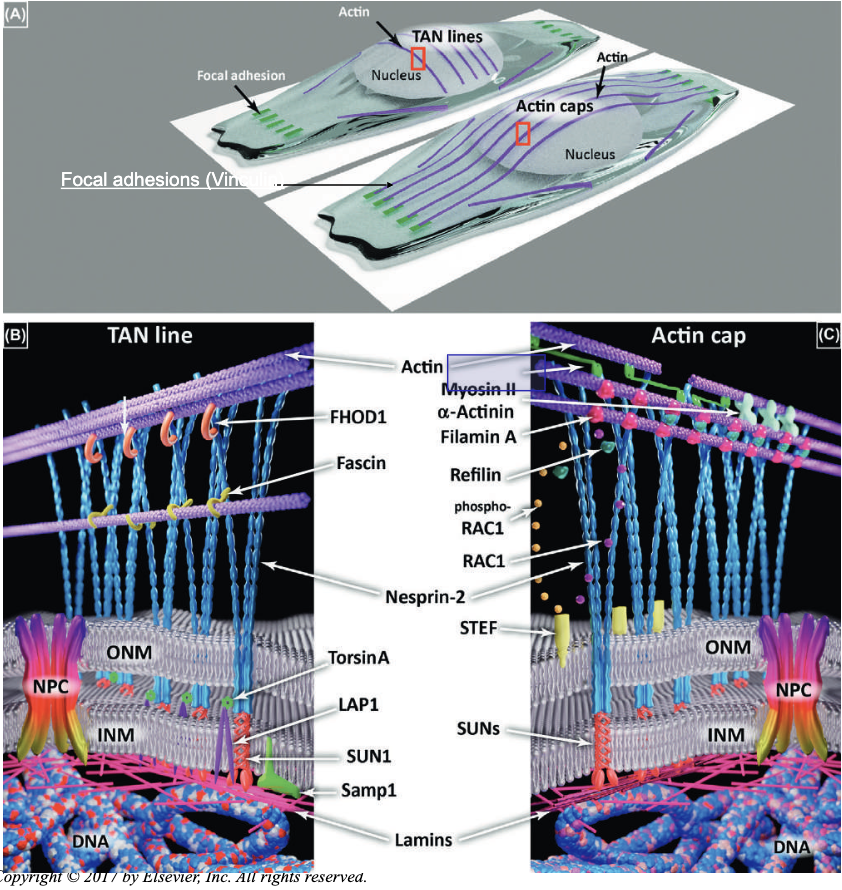
REMEMBER THE SUNS AND THE NESPRINS, SHES NOT GONNA ASK THE OTHERS
Focal adhesions (Vinculin)
- Vinculin:
- cytoskeletal protein associated with cell-cell and cell- matrix junctions.
- It functions as one of several interacting proteins involved in anchoring F-actin to the membrane
- Myosin II:
- non-muscle myosin involved in cell motility, adhesion and cytokinesis → provides contractility to stress fibers.
Proteins will make connections with the NESPRIN: such as the SUNs and actin cap to maintain actin filaments in place, anchoring them to the membrane.
Actin filaments (actin cap) lock the vinculin in shape, squeezing the nucleus in place. These are also useful for movement, so they’re decorated with non-muscle myosin II, permitting contractility.
There are also fibers that lose the myosin connection, and those are the actin fibers in the basal side, underneath the nucleus. Those fibers are not needed for motility, but as a backbone for the general structure.
The gel-like network
The gel-like network organization of actin filaments which form a cross-linked network similar to a gel.
The stiffness is due to the protein filamin, that assists the formation of the actin network forming high-viscosity matrices such as those of the cell cortex
Parallel Bundles
Are thin filaments associated with the proteins fimbrin and villin, which are responsible for forming closely packed parallel bundles of actin that form the core of microspikes (filopodium) and microvilli.
These bundles are anchored to the terminal web - a region of the cortex composed of the network of intermediate filaments and the protein spectrin.
Thin-Filaments in filopodia and lamellopodia in cell migration
filopodia
lamellopodia
Actin in focal contacts
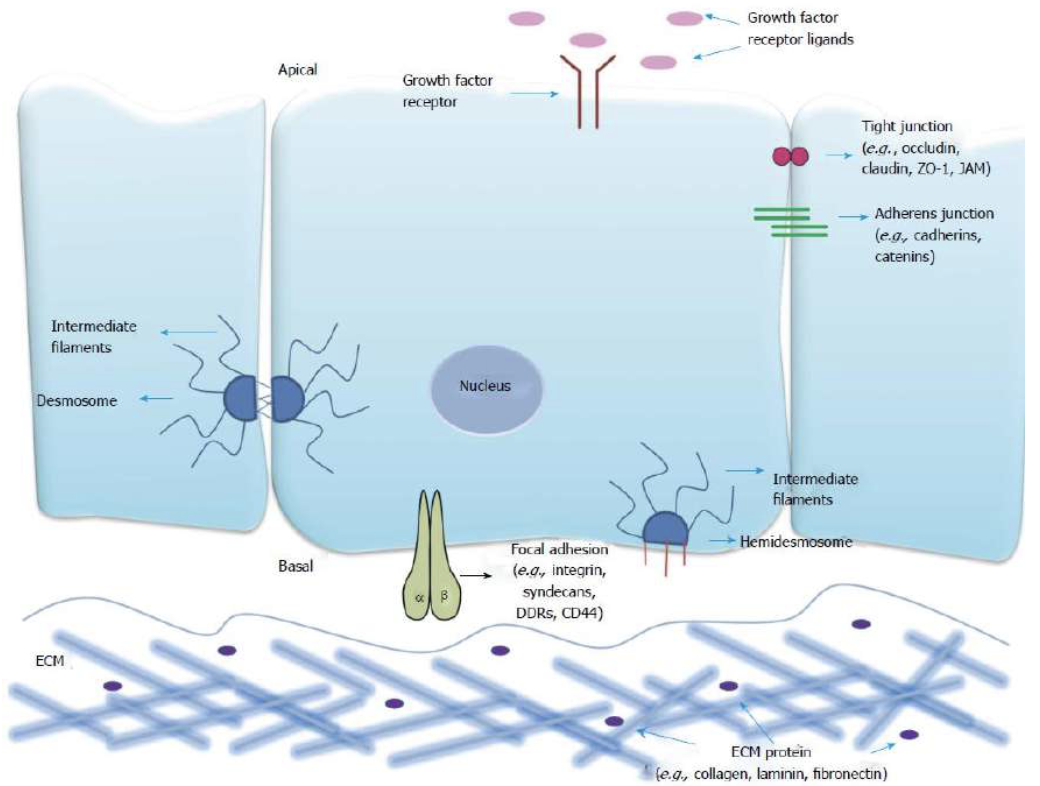
In all cells with an epithelial origin, we always have a substrate where the cells attach to.
When an epithelial cell loses the capability to stay attached to the ECM, it can only undergo apoptosis.
The absence of this reaction is linked to a known cancer pathway.
Along with integrin, actin is important in the establishment of focal contacts of the cells with the extracellular matrix.
The ECM is made of multiple fibers + proteins.
At the focal contacts the transmembrane molecules integrins bind glycoproteins such as fibronectin in the ECM, permitting the cells to maintain the attachment.
The intracellular region of the integrins contact the cytoskeleton and binds the Talin, which contacts both actin filaments and vinculin, forming contractile bundles, known as stress fibers.
Actin doesn’t have direct contact with the integrin, but they do attach to the talin (bridge protein between integrin and actin filament).
All together they provide the strength for this filament to stay attached, and eventually to stretch very strongly.
manca della roba da aggiugnere. organizzare sta ultima roba su actin filopodia bla bla non si capisce un CAZZO
Intermediate filaments
Intermediate filaments and their associated proteins assist in the establishment and manteinance of the three-dimensional framework of the cell.
They are so named because their diameter is intermediate between the smaller actin filaments and the larger microtubules.
These filaments and their associated proteins:
- Provide structural support for the cell
- Form a deformable 3D structural framework for the cell
- Anchor the nucleus in place
- Provide an adaptable connection between the cell membrane and the cytoskeleton
- Maintenance of the nuclear envelope as well as its reorganization subsequent to mitosis
Main function:
- Protect the structural and functional integrity of the cells from external stresses by forcing modulations in the shape and location of cellular constituents.
Intermediate filaments are found in the cells of most animal tissues, where they form a network of fibers that help to maintain the shape and integrity of the cell.
One of the important functions of intermediate filaments is to provide mechanical strength to cells and tissues.
For example,
in epithelial tissues such as the skin, keratin intermediate filaments provide structural support to the cells and help to prevent damage from mechanical stress or abrasion
The Intermediate Filaments are rope-like filaments that are constructed of 8 tetramers of proteins bundled into helics.
Each monomer has N-terminus (head) and C-terminus (tail), both folded into globular domains.
Intermediate filaments are tissue specific.
The categories of intermediate filaments:
- Keratins → specific for epithelial cells
- Desmin → specific of all type of muscle cells
- Vimentin → specific of cells of mesenchymal origin, fibroblast and endothelial cells
- Neurofilaments → specific of neurons
- Nuclear Lamins
qua
c’è una tabella che va molto piu in profondità… tene frega qualcosa…?
EMT (Epithelial Mesenchimal Transition)
When normal epithelial cells turn into cancer, they will produce both keratins but also vimentin. This is called EMT, a wide example of tissue transformation.
Critical Thinking Question: Tumor of Unknown Primary
About 5-10% of cancer patients suffer from cancer of unknown primary (CUP).
The origin of these tumors assists not only in the diagnosis but also in devising effective treatment plans .To recognize the cancer of origin, classifying the types of intermediate filaments present in the cell is crucial.
Microtubules –
- Long, straight and rigid cylindrical structures.
- Provides rigidity and maintains cell shape.
- Function specific to the microtubules: Regulate intracellular movement of organelles and vesicles.
- Establish intracellular compartments.
- Provide the ciliary motion → i.e. sperm cells.
Follows the same organization of Microfilaments.
Monomers of tubulins ( & ) are scattered into the cells, by forming heterodimers.
- The region where the gamma-tubulin ring complex begins the nucleation process is called centrosome
- Centrosome is considered the microtubules organizing center of the cells (MTOC)
- Microtubules are polarized, having a plus end (beta-tubulin) and minus end (alpha tubulin).
- MT are dynamic structures that change their length by undergoing growth burst and then become shorter.
- The minus end depolymerizes, thus shortening the microtubules.
- Alpha/beta tubulin heterodimers are assembled into protofilaments.
Microtubules provide Ciliary Motion
Microtubules make up two types of cellular appendages important for motion:
cilia and flagella.
Cilia are found on many cells of the body, including the epithelial cells that line the airways of the respiratory system.
Cilia move rhythmically; they beat constantly, moving waste materials such as dust, mucus, and bacteria upward through the airways, away from the lungs and toward the mouth.
For example,
Beating cilia on cells in the female fallopian tubes move egg cells from the ovary towards the uterus.
- https://unibo.smartzoom.com/s1241/course1776/f1777/i1789/
- https://unibo.smartzoom.com/s1241/course1776/f1777/i1790/
A flagellum (plural = flagella) is an appendage larger than a cilium and specialized for cell locomotion.
The only flagellated cell in humans is the sperm cell that must propel itself towards female egg cells.
Navigator‐3, a modulator of cell migration
Microtubules are also important to the regulation of cell migration.
NAV3 localizes to the plus ends of microtubules and enhances their polarized growth.
The NAV3 protein augments the ability of cells to adhere to a migration course, while also slowing down migration rates.
Video Analysis:
Co-expression of GFP‐NAV3 and mCherry‐α‐tubulin demonstrated dynamic tracking of the tips of growing MTs by NAV3
Red → Microtubules
NAV3 → Green
NAV3 marks the tips of the growing MT. It’s an indicator of the elongation of the microtubules. This could be for example, a way for the cell to reach and internalize an endosome.
Centrioles
Centrioles are small, cylindrical structures, 0.2 um in diameter and 0.5 um in length.
Centrioles are paired structures, arranged perpendicular to each other and are embedded in a matrix of pericentriolar material (Gamma Tub and pericentrin both interacting with the microtubules and anchoring them to the centrosome).
They are composed of nine triplets of microtubules arranged around a central axis.
Each triplet is arranged so that it forms an oblique angles with the adjacent and a straight angle (90°) with the fifth triplet-
During the S phase of the cell cycle, each centriole of the pair replicates, forming a procentriol in some unknown manner.
Sample Questions
Question
Microfilaments:
a. are 6-7nm thick
b. are composed of two chains of globular subunits
c. all of them
d. are actin filaments that interact with myosin to allow cellular movement
e. possess a faster growing plus end and a slower growing minus endThe answer is C.
Question
The protein filamin associates to:
a. tubulin
b. all of them
c. myosin ii
d. membrane
e. gel-like networksThe answer is E.
Question
Select the wrong answer: contractile bundles are:
a. relevant for cell metabolism
b. relevant for organelles
c. relevant for cell division
d. relevant for cell proliferation
e. relevant for cell migrationThe answer is A.
TARGET DECK: MED::I::Morphology and Development::Histology::02 - Cytoplasm
What are the three main components of the cytoskeleton and their respective diameters?
- Microfilaments (Thin filaments):
- Intermediate filaments:
- Microtubules:
Anki cloze
{1:Treadmilling} is the process where actin monomers are added to the {2:plus (barbed)} end and removed from the {3:minus (pointed)} end.
Anki cloze
Which proteins regulate the assembly and disassembly of actin during treadmilling?
- {1:Profilin}: Binds {2:G-actin} and {2:facilitates addition} to the {2:plus end}.
- {1:Cofilin}: Induces {2:depolymerization} at the {2:minus end}.
- {1:Capping proteins (e.g., Gelsolin)}: Stabilize the {2:plus end} to {2:terminate lengthening}.
Anki cloze
The nuclear envelope is connected to the actin cytoskeleton via the {1:LINC} complex, specifically involving {2:nesprins} and {3:SUN} proteins.
Anki cloze
List the tissue-specific intermediate filaments.
- Epithelial cells: {1:Keratins}
- Muscle cells: {1:Desmin}
- Mesenchymal cells (fibroblasts): {1:Vimentin}
- Neurons: {1:Neurofilaments}
- Nucleus: {1:Nuclear Lamins}
Anki cloze
The {1:Centrosome} acts as the Microtubule Organizing Center (MTOC), where {2:-tubulin} rings nucleate the growth of new microtubules.
What is the structural arrangement of a centriole?
Nine triplets of microtubules arranged in a cylinder, typically found in pairs at right angles () to each other.
Anki cloze
In cell migration, {1:lamellipodia} are broad, sheet-like protrusions, while {2:filopodia} (microspikes) are thin, finger-like projections containing parallel actin bundles.
What is the role of Vinculin and Talin in focal adhesions?
They act as bridge proteins:
- Integrins bind the ECM,
- Talin binds the integrin,
- and Vinculin helps anchor F-actin stress fibers to the complex, providing structural stability and contractility.