TARGET DECK: MED::I::Signaling Pathways in Health and Disease::Metabolic Biochemistry::05 - Glycogen Metabolism
Overview
Glycogen is the body’s carbohydrate “battery” — a rapidly mobilisable reserve that bridges meal-to-meal fluctuations in glucose availability.
Blank container
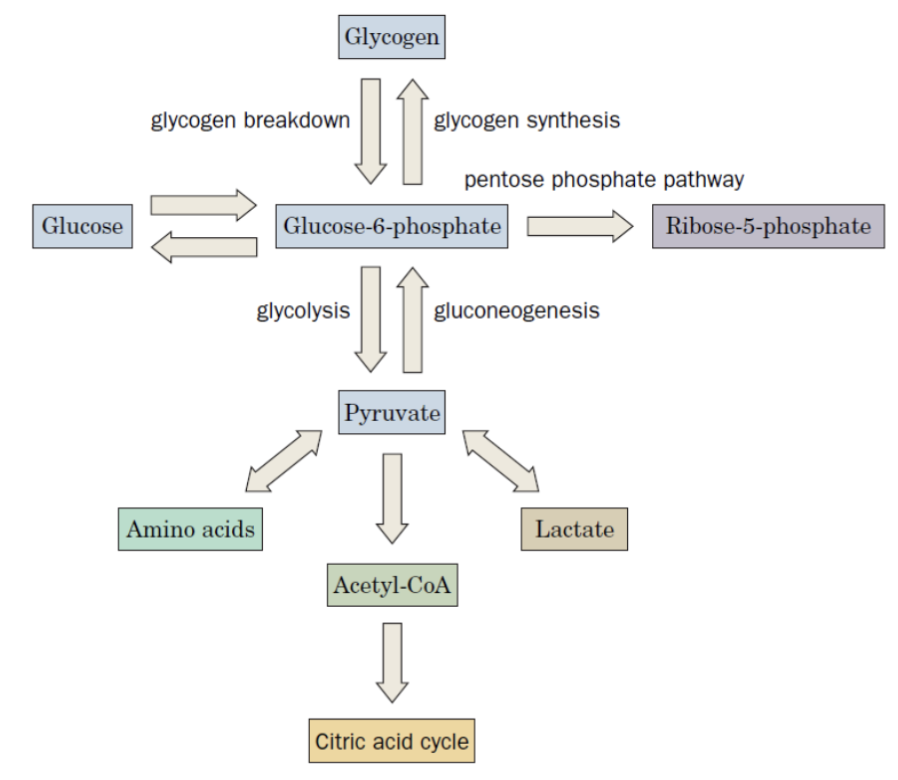
To understand glycogen metabolism, it helps to first see where glycogen sits in the broader metabolic map:
From glucose-6-phosphate, the cell can branch in several directions depending on its needs:
- Glycolysis / Gluconeogenesis → Pyruvate → Amino acids, Lactate, Acetyl-CoA
- Pentose phosphate pathway → Ribose-5-phosphate
The key insight is that glycogen is not just a passive storage depot — its synthesis and breakdown are tightly regulated, tissue-specific, and integrated with the hormonal state of the whole organism.
Tissue-Specific Regulation
Before diving into the biochemistry, it’s worth asking: why does glycogen metabolism need to differ between tissues?
The answer is that the liver and muscle have fundamentally different jobs:
Liver
Controls blood glucose homeostasis — it acts on behalf of the whole organism
The liver releases glucose into the blood when levels drop, and stores it when levels are high. Its glycogen metabolism is therefore driven by hormonal signals reflecting whole-body glucose status.
- Insulin (post-meal) → promotes glycogen synthesis
- Glucagon (fasting) → promotes glycogenolysis
Muscle
Responds to local energy demands — it acts on behalf of itself
Muscle glycogen is never exported as free glucose (muscle lacks glucose-6-phosphatase). It exists purely to fuel contraction.
- High ATP → glycogen synthesis
- Low ATP / contraction → glycogenolysis
This distinction will come up repeatedly — keep it in mind as you read on.
Glycogen Granule Structure
Glycogen isn’t just a loose polymer floating in cytoplasm. In hepatocytes, glycogen exists as organised granules that co-localise with all the machinery needed to regulate it:
- Glycogen polymer
- Synthetic and degradative enzymes
- Regulatory machinery (kinases, phosphatases)
This co-localisation means that regulation can be fast and local.
Glycogen Synthesis (Glycogenesis)
The Core Problem: Activating Glucose
To add glucose to a growing glycogen chain, the cell first needs to make it reactive enough to form a new glycosidic bond. Free glucose isn’t reactive enough — it needs to be activated.
The solution is to attach glucose to UDP, forming UDP-glucose, which acts as a high-energy glucose donor.
Reaction Sequence
Working through it step by step:
Step 1 — Trap glucose in the cell:
Step 2 — Reposition the phosphate:
Step 3 — Activate glucose by attaching it to UDP:
Step 4 — Transfer glucose onto glycogen:
Step 5 — Recycle UDP back to UTP:
Step 6 — Hydrolyse PPi to drive the whole process forward:
Why use UDP-glucose?
The anomeric carbon of glucose is activated by attachment to UMP via a phosphodiester bond.
This provides:
- Irreversibility — PPi hydrolysis drives the reaction forward
- A good leaving group for nucleophilic attack
- A molecular tag marking glucose for glycogen synthesis
Why is UDP-glucose used in glycogen synthesis?
It activates glucose, provides a good leaving group, and makes the reaction effectively irreversible.
05 - Glycogen Metabolism 2026-05-26 17.01.31.excalidraw
⚠ Switch to EXCALIDRAW VIEW in the MORE OPTIONS menu of this document. ⚠ You can decompress Drawing data with the command palette: ‘Decompress current Excalidraw file’. For more info check in plugin settings under ‘Saving’
Excalidraw Data
Text Elements
Link to original
A Note on Nucleotide Recycling
Steps 3 and 5 above involve nucleotide interconversions. Three enzymes handle these broadly across metabolism — worth knowing their names:
| Enzyme | Reaction |
|---|---|
| Adenylate kinase (myokinase) | |
| Nucleoside monophosphokinase (NMPK) | |
| Nucleoside diphosphokinase (NDPK) |
Glycogen Synthase — The Central Enzyme
Glycogen synthase is the enzyme that actually elongates the glycogen chain. A common point of confusion is its name:
Synthase ≠ Synthetase
- Glycogen synthase = glycosyltransferase; transfers glucose from UDP-glucose to glycogen
- Synthetase / Ligase uses ATP to condense two molecules:
Glycogen synthase is also the major regulatory target in glycogenesis — we’ll come back to this in detail.
Why Branching Matters
Glycogen synthase only makes α(1→4) linkages — a linear chain. But linear chains have a problem: they’re relatively insoluble and have only one non-reducing end (the only site where synthesis or breakdown can occur).
Branching enzyme (transglycosylase) solves both problems: it cleaves an α(1→4) bond and reforms it as an α(1→6) bond, creating branch points. The result:
- More soluble polymer
- Exponentially more non-reducing ends → faster synthesis and breakdown
Glycogenin — How Does It All Start?
There’s a chicken-and-egg problem: glycogen synthase needs a glycogen chain to extend — but where does the first chain come from?
The answer is glycogenin, a self-glucosylating protein that acts as the primer:
Glycogenin
- UDP-glucose forms a glycosidic bond with Tyr194 of glycogenin (self-glucosylation)
- Seven more UDP-glucose units are added to build an oligosaccharide
- Glycogen synthase then takes over for chain elongation
Glycogenin remains covalently attached at the core of every mature glycogen granule.
Glycogenolysis
The Core Strategy: Phosphorolysis, Not Hydrolysis
When glycogen needs to be broken down, you might expect simple hydrolysis (adding water). Instead, the cell uses phosphorolysis — adding phosphate instead of water. This is a crucial design choice:
Why? Because phosphorolysis directly produces glucose-1-phosphate rather than free glucose. The phosphate group is added for free (from inorganic phosphate), saving one ATP compared to phosphorylating free glucose later.
Reaction Sequence
The last step only occurs in the liver. Muscle cannot export free glucose — glucose-6-P is committed to glycolysis.
The Branch Problem
Glycogen phosphorylase can only cleave α(1→4) bonds — it has no way to deal with branch points. Worse, it stalls 4 residues from a branch point and can go no further.
Phosphorylase limitation
Glycogen phosphorylase stalls 4 residues from a branch point. Debranching enzyme must act first.
How far from a branch point does glycogen phosphorylase stall?
Four residues from the branch point.
This is where the debranching enzyme comes in. It has two distinct activities working in sequence:
- Transferase activity: moves the trisaccharide “stub” from the branch to the main chain (α1→4 → α1→4), exposing the single α(1→6)-linked glucose
- Glucosidase activity: hydrolyses that last α(1→6) glucose → releases it as free glucose (not glucose-1-P)
What are the two activities of the glycogen debranching enzyme?
Transferase activity moves a trisaccharide to the main chain, and glucosidase activity removes the remaining α(1→6)-linked glucose.
Note that this one free glucose per branch is the only glucose released by glycogenolysis that isn’t phosphorylated — it bypasses the need for hexokinase but also can’t directly feed glycolysis without re-phosphorylation.
Glycogen Phosphorylase — Regulation in Detail
Glycogen phosphorylase is the rate-limiting enzyme of glycogenolysis and is regulated at multiple levels simultaneously. Understanding its regulation is central to understanding the whole section.
The Two Forms
Phosphorylase exists in two interconvertible forms, controlled by phosphorylation:
| Form | Ser14 | Activity |
|---|---|---|
| Phosphorylase a | Phosphorylated | Active |
| Phosphorylase b | Unphosphorylated | Less active |
The interconversion is catalysed by:
Additional Regulatory Sites
But phosphorylation isn’t the whole story. Phosphorylase also has allosteric sites that allow it to respond to the cell’s immediate energy status:
| Site | Ligand / Modification | Effect |
|---|---|---|
| Active site | Glycogen + | Catalysis |
| Allosteric site | AMP | Activation |
| Allosteric site | ATP, Glucose | Inhibition |
| Phosphorylation site (Ser14) | Phosphorylated (form a) | Active |
| Phosphorylation site (Ser14) | Unphosphorylated (form b) | Less active |
| Cofactor | Pyridoxal-5’-phosphate (PLP) | Required for catalysis |
Tissue-specific regulatory dominance
The same enzyme is regulated differently depending on the tissue’s role:
- Muscle: allosteric regulation (ATP/AMP) dominates → responds to local energy charge within seconds
- Liver: phosphorylation dominates → responds to hormonal signals reflecting whole-body glucose
A Clever Feedback Loop in the Liver
Once blood glucose rises (e.g. after a meal), the liver needs to stop breaking down glycogen — even before hormonal signals can reverse the phosphorylation cascade. Glucose itself provides a direct brake:
Glucose feedback in liver
Glucose binds an allosteric site on phosphorylase a → conformational change exposes Ser14 to PP1 → dephosphorylation → inactivation.
This auto-brakes glycogenolysis when blood glucose rises, before insulin even acts.
The Hormonal Activation Cascade
When blood glucose is low, glucagon (liver) or epinephrine (liver + muscle) trigger glycogenolysis through a phosphorylation cascade:
Why use a cascade?
Each enzymatic step amplifies the signal: one hormone receptor → many cAMP molecules → many PKA molecules → many phosphorylase kinase molecules → many phosphorylase a molecules → rapid, massive glycogen breakdown.
This cascade achieves both speed and amplification from a tiny hormonal stimulus.
How PKA Is Activated by cAMP
Inactive PKA is a tetramer (2 regulatory + 2 catalytic subunits). The regulatory subunits hold the catalytic subunits inactive.
cAMP binds the regulatory subunits → they release the catalytic subunits → catalytic subunits are now free and active.
Signal termination: cAMP phosphodiesterase (PDE) hydrolyses cAMP → 5’-AMP. (PDE can itself be activated by PKA — a negative feedback loop.)
Glycogen Synthase Regulation
Now let’s look at the other side of the equation. Glycogen synthase regulation mirrors phosphorylase — but in reverse:
| Form | Phosphorylation | Activity |
|---|---|---|
| GS-a | Unphosphorylated | Active |
| GS-b | Phosphorylated | Inactive |
Notice that phosphorylation activates phosphorylase but inactivates synthase. This means a single hormonal event (triggering phosphorylation) simultaneously turns on breakdown and turns off synthesis — an elegant and efficient design.
Key kinase: GSK3 (Glycogen Synthase Kinase 3) phosphorylates → inactivates GS.
Insulin counter-acts this:
Insulin → activates PKB (Akt) → PKB phosphorylates and inactivates GSK3 → GS remains active → glycogen synthesis proceeds.
PP1 dephosphorylates both GS (activating it) and glycogen phosphorylase (inactivating it) — the same phosphatase coordinates both enzymes.
Which glycogen synthase form is active?
GS-a, the unphosphorylated form.
Coordination of Glycogen Metabolism
The Core Principle: Reciprocal Regulation
The most important concept in this entire section is reciprocal regulation:
Reciprocal regulation
Phosphorylase and synthase are always regulated in opposite directions — the cell never wastes energy running synthesis and breakdown simultaneously.
This is enforced at multiple levels (allosteric, phosphorylation, and hormonal) and is the reason glycogen metabolism is so efficient.
| State | Phosphorylase | Glycogen Synthase |
|---|---|---|
| Phosphorylated | Active (a) | Inactive (b) |
| Dephosphorylated | Less active (b) | Active (a) |
What is the logic of reciprocal regulation in glycogen metabolism?
Glycogen synthase and glycogen phosphorylase are regulated in opposite directions so synthesis and breakdown do not occur at the same time.
Liver: Hormonal Integration
Putting it all together for the liver:
| Hormone | Phosphorylase | Glycogen Synthase | Net Effect |
|---|---|---|---|
| Insulin | Dephosphorylated → inactive | Dephosphorylated → active | Glycogen synthesis |
| Glucagon | Phosphorylated → active | Phosphorylated → inactive | Glycogen breakdown |
Glucagon’s effects extend far beyond glycogen — it coordinates a whole-body switch to fasting metabolism:
| Metabolic effect | Target enzyme |
|---|---|
| ↑ Glycogenolysis | Glycogen phosphorylase |
| ↓ Glycogen synthesis | Glycogen synthase |
| ↓ Glycolysis | PFK-1 |
| ↑ Gluconeogenesis | FBPase-2, PEP carboxykinase, pyruvate kinase |
| ↑ Fatty acid mobilization | Hormone-sensitive lipase |
| ↑ Ketogenesis | Acetyl-CoA carboxylase |
Glucose as a Direct Signal in the Liver
Hormones aren’t the only regulators. High blood glucose itself acts directly:
High blood glucose → enters hepatocytes (GLUT2) → glucose-6-P accumulates → allosterically activates GS and inhibits phosphorylase.
This provides a fast, hormone-independent way for the liver to sense and respond to glucose.
Glycogen Metabolism in Muscle
Muscle glycogen metabolism follows the same basic biochemistry but with key differences that reflect muscle’s different role:
Key differences from liver
- Glucose uptake via GLUT4 (insulin-dependent)
- Phosphorylation by low-Km hexokinase
- No glucose-6-phosphatase → glucose-6-P cannot leave as free glucose; it feeds glycolysis only
- Regulation dominated by ATP/AMP ratio and Ca²⁺, not blood glucose
How Muscle Matches Glycogenolysis to Contraction
The beauty of muscle regulation is that it’s automatic — glycogen breakdown accelerates precisely when and because contraction is occurring:
- At rest: high ATP → glycogen synthesis
- During contraction:
- AMP accumulates → allosterically activates phosphorylase b (no phosphorylation needed — immediate)
- Ca²⁺ (the same signal that triggers contraction) activates phosphorylase kinase via its calmodulin subunit → generates phosphorylase a (sustained activation)
- GS is simultaneously phosphorylated → inactive
- Glucose-6-P flows exclusively into glycolysis
ATP Production in Skeletal Muscle
Glycogen is not the only ATP buffer in muscle. During intense exercise, the cell draws on several systems in sequence, from fastest to slowest:
Rapid ATP Recovery Systems
This reaction also generates AMP — which then allosterically activates phosphorylase b, linking the energy-sensing and glycogenolytic systems.
Creatine kinase directionality
The same enzyme runs in opposite directions depending on energy state:
At rest (high ATP): (stores energy)
During contraction (high ADP): (releases energy)
Creatine:
OC(=O)CN(C)C(=N)NPhosphocreatine:
OC(=O)CN(C)C(=N)NP(=O)(O)OSequential Activation During Exercise
These systems activate in sequence as exercise continues:
- Immediate (seconds): phosphocreatine buffer
- Early (seconds–minutes): AMP → allosteric phosphorylase b activation
- Sustained: Ca²⁺ → phosphorylase kinase → phosphorylase a
- Hormonal (epinephrine): cAMP → PKA → full cascade
Physiological Effects of Epinephrine
Epinephrine coordinates the whole organism for “fight or flight”:
| Effect | Metabolic Consequence |
|---|---|
| ↑ Heart rate, blood pressure | Increased O₂ delivery to tissues |
| ↑ Glycogenolysis (muscle + liver) | More glucose for fuel |
| ↓ Glycogen synthesis | Glucose not stored |
| ↑ Gluconeogenesis (liver) | Sustained glucose supply |
| ↑ Glycolysis (muscle) | More ATP for contraction |
| ↑ Fatty acid mobilization | Alternative fuel |
| ↑ Glucagon / ↓ Insulin | Reinforces metabolic effects |
Glycogen Storage Diseases
When enzymes in this pathway are defective, glycogen either accumulates abnormally or can’t be mobilised, causing tissue damage. The pattern of symptoms tells you which enzyme is affected and in which tissue:
Clinical relevance
Defects in glycogen metabolism enzymes cause accumulation of abnormal glycogen or hypoglycemia, with tissue-specific consequences.
| Type | Name | Enzyme Defect | Organ | Key Symptoms |
|---|---|---|---|---|
| 0 | — | Glycogen synthase | Liver | Hypoglycemia, high ketones, early death |
| Ia | Von Gierke | Glucose-6-phosphatase | Liver | Hepatomegaly, kidney failure |
| Ib | — | Microsomal G6P translocase | Liver | As Ia + susceptibility to infections |
| II | Pompe | Lysosomal glucosidase | Skeletal + cardiac muscle | Infantile: death by age 2; adult: muscular dystrophy-like |
| IIIa | Cori / Forbes | Debranching enzyme | Liver, skeletal + cardiac muscle | Hepatomegaly, myopathy |
| IV | Andersen | Branching enzyme | Liver, skeletal muscle | Hepatosplenomegaly, myoglobinuria |
| V | McArdle | Muscle phosphorylase | Skeletal muscle | Exercise cramps, myoglobinuria |
| VI | Hers | Liver phosphorylase | Liver | Hepatomegaly |
| VII | Tarui | Muscle PFK-1 | Muscle, erythrocytes | As V + hemolytic anemia |
| VIII/IX | — | Phosphorylase kinase | Liver, leukocytes, muscle | Hepatomegaly |
| XI | Fanconi-Bickel | GLUT2 | Liver | Failure to thrive, rickets, kidney dysfunction |
Exam-Type Question
Q: Reaction proceeds left-to-right:
a. During muscle contraction
b. When the respiratory chain is uncoupled
c. In muscle at rest
d. If creatine kinase content is increased
Answer: C — at rest
- At rest, ATP is abundant → stored as phosphocreatine (left-to-right ✓)
- (a) During contraction, phosphocreatine donates phosphate to ADP → reaction runs right-to-left
- (b) Uncoupling = no ATP formed → ADP accumulates → reaction runs right-to-left
- (d) Enzyme amount never changes reaction direction, only rate
TLDR - 05 - Glycogen Metabolism
Glycogen is a highly branched glucose polymer that serves as the primary carbohydrate reserve — in the liver for blood glucose homeostasis, and in muscle for rapid ATP production.
Synthesis (Glycogenesis)
Glucose is activated as UDP-glucose (irreversible, PPi hydrolysis drives it), then transferred onto glycogen by glycogen synthase (α1→4 bonds). Branching enzyme introduces α1→6 branch points. A protein primer, glycogenin, initiates the chain at Tyr194.
Breakdown (Glycogenolysis)
Glycogen phosphorylase cleaves α1→4 bonds phosphorolytically (releasing glucose-1-P, not free glucose). It stalls 4 residues from branch points; debranching enzyme (transferase + glucosidase activities) resolves branches and releases one free glucose per branch.
Regulation — the key principle
The two key enzymes are always regulated reciprocally:
| State | Phosphorylase | Glycogen Synthase |
|---|---|---|
| Phosphorylated | Active (a) | Inactive (b) |
| Dephosphorylated | Less active (b) | Active (a) |
Hormonal cascade: Glucagon/Epinephrine → adenylate cyclase → cAMP → PKA → phosphorylase b kinase → phosphorylase a (active) + GS-b (inactive) → glycogenolysis.
Insulin reverses this: activates PKB → inactivates GSK3 → GS stays active; activates PP1 → dephosphorylates both enzymes → glycogen synthesis.
Tissue differences
| Liver | Muscle | |
|---|---|---|
| Primary role | Blood glucose buffer | Local ATP supply |
| Glucose transporter | GLUT2 | GLUT4 |
| Glucose-6-phosphatase | ✅ (releases free glucose) | ❌ (glucose-6-P → glycolysis only) |
| Dominant regulator | Hormonal (glucagon/insulin) | Energy charge (ATP/AMP, Ca²⁺) |
ATP buffers in muscle (fastest to slowest)
- Phosphocreatine → immediate (seconds)
- Glycogenolysis → allosteric (AMP) + Ca²⁺ → phosphorylase a
- Epinephrine cascade → cAMP → PKA → full hormonal activation
Glycogen storage diseases
Enzyme defects in the glycogen metabolism pathway cause accumulation of abnormal glycogen or hypoglycemia. Key examples: Von Gierke (no G6Pase → hepatomegaly + hypoglycemia), Pompe (no lysosomal glucosidase → cardiomyopathy), McArdle (no muscle phosphorylase → exercise intolerance + myoglobinuria).