Gallery
General features of cell signal transduction
Blank
Biosignaling
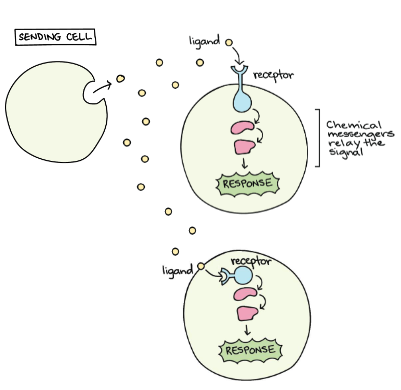
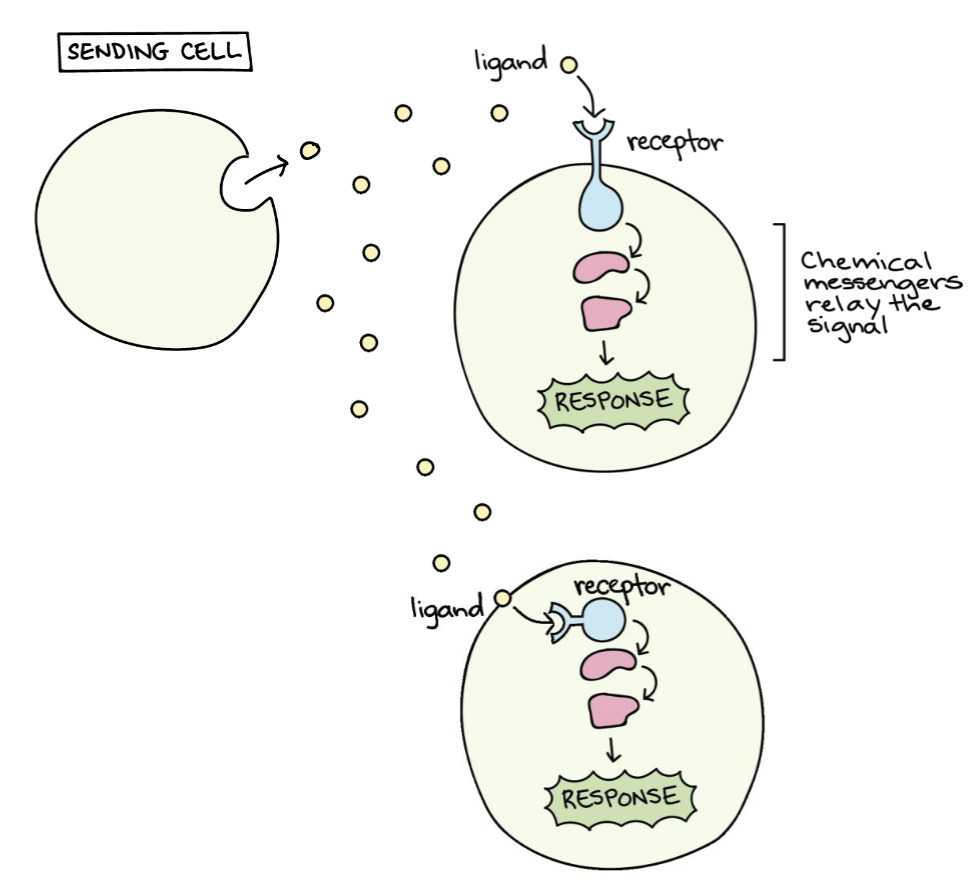
Biosignaling is the ability of cells to convert environmental signals (arriving from the outside) into chemical modifications within the cell.
Extracellular signaling molecules have crucial roles in development and homeostasis, and their incorrect deployment can lead to developmental defects and disease states.
Signaling molecules are released from sending cells, travel to target cells and act over length scales of several orders of magnitude, from morphogen-mediated patterning of small developmental fields to hormonal signaling throughout the organism.
Cells need to communicate with each other.
Key Terms
Every target cell needs to be responsive to a signal, and to have a specific receptor for that signal, which is also called ligand (because it binds to the receptor).
The binding of the ligand to the receptor results in an altered shape of the receptor (because of its proteic nature).
This conformational change will eventually lead to the desired response inside the target cell.
Remember that not all cells can sense a particular message: in the same environment, the same signal is recognized by a target cell that has a specific receptor, while it is not recognized by a cell that does not have the receptor for that particular ligand (some proteins are synthesized only in some cells, depending on chromatin conformation).
Signals are often relayed inside the cell.
Basically, extracellular signals are converted in intracellular signals that are often related through a chain of chemical reactions: this pathway of different chemical steps leads to a final response (alterations in gene expression, metabolic pathways etc.).
All kinds of signals (even physical signals) are converted to chemical signals by the receptor; e. g. photoreceptors and pressure- sensing channels.
Ligands come in many different varieties and interact with the receptor either from the surface of the cell, or from the inside.
Receptor location
Blank
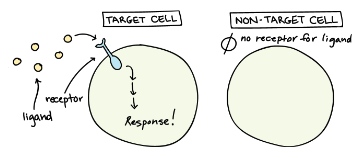
Receptor location is important:
a receptor can either be placed in the cytoplasm or on the cell membrane.
- Receptor on the cell membrane:
- the receptor has a domain that faces the extracellular environment and interacts with the ligand; and an intracellular domain that is able to activate the cascade of signals.
- Receptors in the cytoplasm:
- they can respond only to ligands that can cross the cell membrane (mostly hydrophobic compounds).
If a hormone type is destined for a membrane receptor then it will be received by those kinds of cell (only if they can produce that receptor).
The original extracellular signal is converted into an intracellular signal that triggers a response.
The receptor’s two-domain nature allows the cell to regulate the binding of ligand and the effect of ligand independently.
Expression of a receptor that is not normally expressed in a cell is often sufficient to confer responsiveness to that receptor’s ligand.
This responsiveness often occurs because the cell already expresses the other components necessary for propagating the intracellular signal from the receptor.
Receptors accelerate functions so they can be analogs to enzymes and catalysts.
Signaling Classification
There are four basic categories of chemical signaling found in multicellular organisms: paracrine signaling, autocrine signaling, endocrine signaling, and signaling by direct contact.
The main difference between the different categories of signaling is the distance that the signal travels through the organism to reach the target cell.
This difference is not important at a molecular level but is more significant at a physiological one.
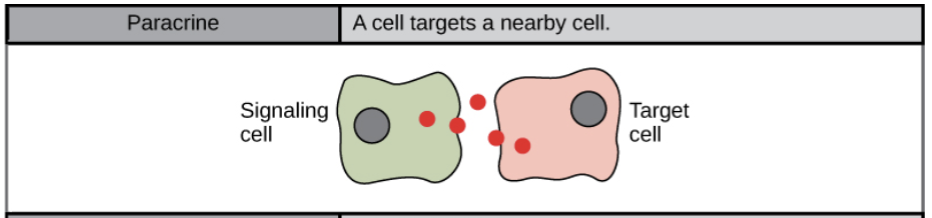
- Paracrine signal:
- often, cells that are near one another communicate through the release of chemical messengers (ligands that can diffuse through the space between the cells). Cells communicate over relatively short distance (immediate surrounding area) to locally coordinate activities with nearby target cells. These signals are especially important during development, when they allow one group of cells to tell a neighboring group of cells what cellular identity to take on.
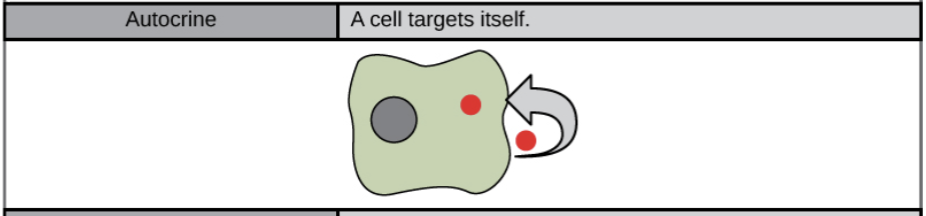
- Autocrine signal:
- a cell signals to itself, releasing a ligand that binds to receptors on its own surface (or, depending on the type of signal, to receptors inside of the cell). For instance, autocrine signaling is important during development, helping cells take on and reinforce their correct identities. From a medical standpoint, autocrine signaling is important in cancer and is thought to play a key role in metastasis (the spread of cancer from its original site to other parts of the body). Usually it is used for negative feedback.
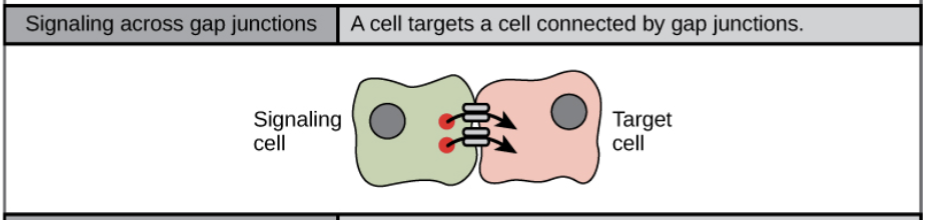
- Cell-cell contact:
- small signaling molecules called intracellular mediators diffuse through neighboring cells that are connected through gap junctions, that are tiny water-filled channels that directly connect the two neighboring cells (small molecules, such as calcium ions, are able to move between cells, but large molecules like proteins and DNA cannot fit through the channels without special assistance). This allows a group of cells to coordinate their response to a signal that only one of them received.
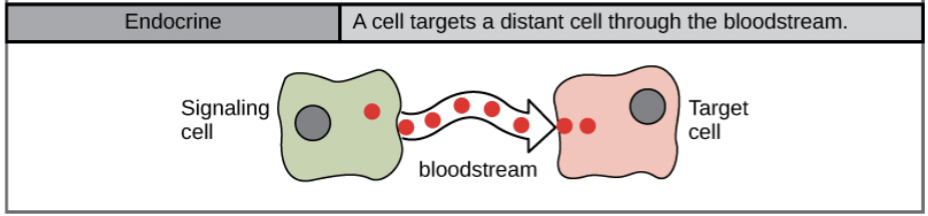
- Endocrine signal:
- the molecules travel a long distance to reach the target. When cells need to transmit signals over long distances, they often use the circulatory system as a distribution network for the messages they send. In long-distance endocrine signaling, signals are produced by specialized cells and released into the bloodstream, which carries them to target cells in distant parts of the body. Signals that are produced in one part of the body and travel through the circulation to reach far-away targets are known as hormones.
Principles of Ligand-Receptor Interactions
- First, the signaling range can be influenced by the amount of the signal that is produced;
- High rates of signal production result in high levels of extracellular signal.
- Second, the strength or signaling ability of a ligand affects its signaling range.
- For example, a mutation or polymorphism that decreases receptor binding but does not affect the distribution of a ligand will nonetheless shorten its signaling range.
- Third, the ability of a signal to move through a tissue will affect its signaling range;
- Molecules that move more freely or directionally through tissues will move farther from their source than molecules that are restricted in their movement.
- Fourth, signal stability helps determine signaling range.
- For example, very stable signals can move a long distance away from their source before being degraded.
- Finally, even highly expressed, stable, active and diffusive signals can have short-range effects because of dilution in target tissues.
Specificity and affinity, dissociation constant. The same theory can be applied to hormones and receptors and signals and receptor. Is a mater of possibility of interaction or not. The receptor won’t be able to transform the signals: in most of the cases the receptor binds and interact with the signals but is not able to transform the structure of the signal. The shape of the receptor is able to recognize the shape of the ligand. The receptor is a transmembrane protein, intracellular part that is protruding in the cytosol -> color code means something. Something is different. There are molecular differences.
The same hormone can have different outcomes within the cell depending on the entire structure of the receptor: also the inner part of the receptor that makes a difference.
The two receptors are identical for what concern the inner parts of the cell but we have a different shape and structure of the parts of the receptor that binds to different ligands (specificity) -> two different messages are interpreted into the same outcome.
Thymeric receptor -> you create it.
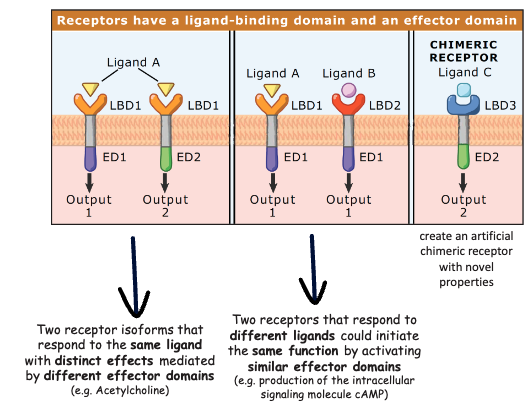
- Specificity:
- Precise molecular complementarity between the signal molecule and its receptor (it can be weak, reversible, non-covalent forces)
- Affinity:
- The same of enzyme-substrate complexes
- Kd=conc of ligand multiplied by concentration of receptors divided by concentration of ligand enzyme complexes
Important
If the is lower than , it means the dissociation constant is very low and therefore the receptor and ligand have high affinity.
Remember that the Kd is the opposite of the constant of association.
- Saturation:
- Like for enzymes, the saturation curve of receptor-ligand is a hyperbolic curve that loses the direct proportion. The receptor is limited so even adding more and more ligand the receptor is already at its own threshold.
- Receptors follow a Michaelis Mentel behavior like enzymes do and it explains the efficiency of the receptor-ligand affinity.
- Cooperativity:
- small changes in ligand concentration can create large changes in receptor activation. The binding of two (or more) similar molecules increases/decreases the receptor affinity for that molecule (/those molecules). All the protein system of a receptor can adapt to the first ligand and become more prone to activate other protein receptors so that the signal will be higher thanks to a positive modulation (further enhancement).
Receptor States: Active and Inactive Form
Receptors can exist in inactive () or active () molecular conformations. The two forms are in equilibrium but we expect that once ligand is not present most of the receptors must be inactive. The equilibrium of these two forms is . In conditions of lack of ligand is mostly towards the left (inactive form).
Ligand binding changes the receptor conformation and L can bind to the receptor in either of its conformations.
We cannot exclude that some receptors remain active even in lack of ligand, therefore once ligand is added we will have both inactive R-L complexes and already active R-L complexes. Soon after inactive R will turn in its active form.
→ Ligands that bind with higher affinity for the conformation than for will be activators (agonists, )
→ Thus, if binding to the RL configuration is preferred → , then ligand binding to any R will shift the conformation equilibrium of RL to the RL state to an equivalent extent → J*/J>>1
For the time that ligand is in the receptor, the receptor will adapt its shape to that of the ligand. The ligand can be released through a change of conformation of the recepetor itself or throguh allosteric modulators for example
If is always active it is a problem because it means that it’s not controlled and cannot be manipulated anymore (cancer)
Info
→ affinity constant
→ equilibrium constant
→ ligand concentration
Important
Ligand Activity:
- Agonists: Bind preferentially to the R* (active) state, shifting the equilibrium toward activation.
- Antagonists: Bind equally to R and R*, blocking the site without changing the equilibrium.
- Inverse Agonists: Bind preferentially to the R (inactive) state, reducing even the basal activity of the receptor.
Multistep signaling pathways
Properties of Receptors
J must be always greater than 0 thus, even unliganded receptors have some activity (R*).
Because physiological receptors are nearly inactive in the absence of ligand, J must be much less than 1 and is probably less than 0.01; most receptors are less than 1% active (R*) without agonist.
Chemical manipulation of a ligand’s structure varies its selectivity between R and R* and can often alter its activity as an agonist. Overexpressed receptors frequently display their intrinsic low activity
A ligand that binds equally well to both the R and R* states will not cause activation, but may still occupy the binding site and thereby competitively inhibit binding of an activating ligand. Such competitive inhibitors (antagonists) are frequently used as drugs to block unwanted activation of a receptor in various disease states
A ligand that binds preferentially to R relative to R* will further shift the conformational equilibrium to the inactive state and cause net inhibition (inverse agonist) In this case, K/K*>>1 → J/J*>>1 BUT given that J<<<J<<1
Amplification
The RL complex activates an enzyme that catalyzes the activation of many molecules and so on in a cascade
Integration
Multiple signals with positive or negative sign are integrated to modulate a unified response
Desensitization
A receptor could me modulated allosterically through feedback circuits
When a signal is present continuously, R no longer responds until the stimulus falls below a certain threshold
During this time, called refractive period, the cell is not able to respond to signals, then sensitivity slowly returns to normal.
Important
Core Signal Processing:
- Amplification: One signal molecule triggers a cascade activating thousands of downstream molecules.
- Integration: The cell “sums up” multiple inputs to create a single coordinated response.
- Desensitization: Prevents overstimulation; the cell enters a refractive period where it stops responding to a continuous signal.